
Пузырчатая головня. Описание, симптомы, источники инфекции, меры контроля и средства защиты растений от болезни представлены на сайте «Сингента» в России.
Признаки заражения
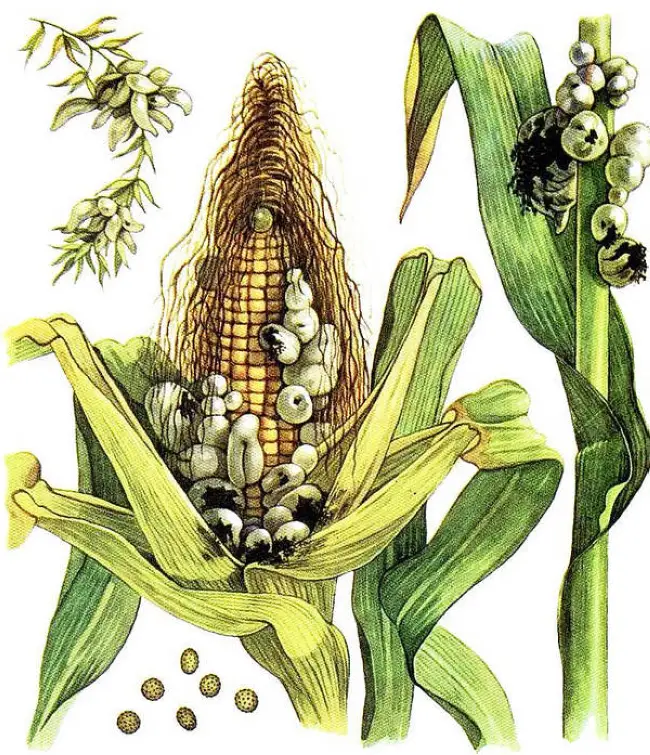
Распознать это заболевание кукурузы не составит никакого труда. Главным симптомом этого заболевания является образование желваков, вздутий различной формы и величины, которые обнаруживают на:
- початках;
- соцветиях;
- стеблях;
- листьях;
- почках.
После заражения головневым грибком ткани растения начинают припухать, меняя свой цвет на более светлый. После чего в этих местах образуется клубневидное или шарообразное вздутие зеленоватого или желтого цвета, заполненное слизью. Через пару недель новообразование сначала темнеет, а затем высыхает и приобретает светло-зеленый или оливковый оттенок. В этом сухом пузыре находятся грибные телоспоры.
После полного созревания тело грибницы становится темного, почти черного цвета. Далее это вздутие лопается, рассыпается и телоспоры разносятся ветром, заражая здоровые растения и грунт.
Зараженный грунт может послужить первичным источником заражения здоровых всходов молодой кукурузы.
При поражении листвы, вздутия имеют вытянутую, продолговатую форму, и образовываются вдоль прожилок. На початках кукурузы поражаются отдельные завязи, которые после заражения останавливаются или задерживаются в росте. На мужском соцветии или султане чаще всего поражаются отдельные цветки, которые образуют пузыри.
Самые крупные вздутия наблюдают в основном на почках и стеблях кукурузы. На листьях они имеют более маленькие размеры, выглядят как шершавая морщина и высыхают еще до созревания телоспор.
Профилактика пузырчатой головни кукурузы

К профилактическим мерам, предупреждающим возникновение пузырчатой головни, относят:
- использование здорового семенного материала,
- применение устойчивых к заболеванию сортов кукурузы,
- высев кукурузы в прогретую землю,
- глубокая зяблевая вспашка почвы осенью.
Сложность природы полевой устойчивости к болезни, трудности создания универсальных инфекционных фонов и необходимость многолетнего изучения селекционного материала в различных экологических условиях привели выводу о разделении полевой устойчивости к возбудителю пузырчатой головни на структурную и физиологическую. Методы искусственного заражения кукурузы U. maydis неоправданно широко распространились в системе НИИ и Государственного сортоиспытания благодаря возможности получать с их помощью высокий уровень пораженности стеблей и початков, практически не наблюдаемый в природе, поскольку негативный отбор позволяет выбраковать восприимчивые формы в процессе селекции исходного материала и многолетних испытаний. Устранение при скрининге структурной устойчивости початков, даже при введении суспензии U. maydis в зону над метелкой и початком без их травмирования, привело к выявлению образцов с физиологической устойчивостью (с минимальным размером галлов). При этом различия в распространенности болезни, обусловленные продолжительностью периода восприимчивости, стали редко адекватны природным, связанным со спецификой повреждения растений комплексом вредителей. Ускользала от внимания исследователей избирательность как тип устойчивости растений к насекомым, её влияние на репродуктивный потенциал вредителей, учитываемые при отборе на групповую устойчивость к вредным организмам. Безусловно, технологические преимущества в воссоздании эпифитотийного уровня развития болезни при искусственной инокуляции очевидны, но в жертву технологичности приносятся особенности и степень выраженности структурного иммунитета, которые при искусственной инокуляции нивелируются, сужая возможности отбора. И одна из важных задач исследований состоит в использовании этого природного фона первичной дифференциации (осуществляемой фитофагами) для выявления наименее повреждаемых и поражаемых селекционных образцов.
Многолетние исследования и анализ литературы привели нас к заключению, что, в отличие от патосистем многих облигатных паразитов, патосистема Zea mays — Ustilago maydis характеризуется рядом следующих биологических особенностей:
- возбудитель в большей мере способен самостоятельно проникать в ткани растения-хозяина (корни, мезокотиле, стебли, листья, початки, пазушные почки) и вызывать их первичную колонизацию в биотрофной стадии жизненного цикла, чем завершать в нем цикл развития формированием галла, что обусловлено особенностями морфо-анатомической защиты — плотным укрытием конуса нарастания и других меристем, преимущественно недоступных для патогена при ненарушенной структурной целостности органов и тканей растения;
- длительный период формирования различных органов растений и восприимчивых к заражению меристем в онтогенезе, продолжительный латентный период, связанный с патологическим перерождением меристем, формированием галлов и созреванием телиоспор, — определяют развитие одного поколения гриба на растении, отмечаемого в онтогенезе в разное время (при формировании ранней и средней листовой воронки, при выдвижении метелок и початков, в период налива и созревания зерна);
- высказываемая представителями ряда фирм мысль о развитии в природных условиях так называемых 3-5 генераций гриба за вегетацию кукурузы — лишь предположение. Видимость нескольких генераций гриба может сложиться без учета влияния разных инвазионных фонов: шведских мух, способствующих формированию и наиболее раннему проявлению галлов головни; первого поколения кукурузного мотылька — появления второй “волны” растений, пораженных U. maydis; второго поколения кукурузного мотылька и хлопковой совки — третьей “волны” пораженных растений. Растянутость лета вредителей, сроков повреждений и продолжительности питания определяют и длительный период проявления головни, что создает видимость развития нескольких генераций U. maydis. Случаи развития на растении второй генерации U. maydis крайне редки, они возможны лишь при переносе инфекционного начала с галлов, созревших до цветения в раневые каналы метелок, основного и рудиментарных початков, преимущественно у линий средней и поздней групп спелости. Две-три генерации телиоспор можно получить лишь при искусственном введении инокулюма к меристемам, используя первоначально вакуум-метод заражения проростков, или в коллекционных питомниках, представленных образцами разных групп спелости, но практически невозможно развитие двух поколений в посевах конкретного гибрида.
В серии публикаций (Иващенко, 1992, 2003; Иващенко и др., 2008; Иващенко, 2009, 2010) нами показано, что паразитоценоз кукурузы формируется в тесной связи с комплексом вредящих фаз фитофагов (шведская муха, кукурузный мотылек, хлопковая совка), первоначально создающих условия для внедрения грибной инфекции и колонизации ими меристем, что обусловлено эволюционно сложившейся общностью пищевых предпочтений группы фитофагов и U. maydis (тропностью), онтогенетической сопряженностью развития фитофагов и растений, что облегчает, надо полагать, реализацию агрессивности возбудителя болезни. В отличие от имеющих место стохастических связей (например, Oscinella frit — Sorosporium. reilianum), возбудителя пузырчатой головни и кукурузного мотылька характеризует стабильность консортивных межпопуляционных связей, их весьма положительная сопряженность (по Уранову, 1935) или зависимость (по Василевич, 1972), при которой увеличение численности фитофага вызывает увеличение численности патогена, что служит основанием для уточнения расчетов суммарной вредоносности фитофагов и патогенов, установления её пороговых значений, разработки прогноза распространения болезней. Кроме того, рассмотрение паразитоценоза кукурузы на уровне тритрофа в большей мере раскрывает возможности использования энтомофагов (трихограмма, габробракон и др.), их роль в снижении численности популяций не только фитофагов, но также распространения пузырчатой головни. Нам представляется (Воронин и др., 1999), что необходима и реальна дальнейшая интеграция иммунитета растений и биометода как биоценологической основы стратегии совершенствования фитосанитарных технологий в агроэкосистемах, призванной обеспечить оптимально-устойчивый биоценотически саморегулирующийся фитосанитарный режим.
При характеристике входящего в паразитоценоз облигатного патогена U. maydis необходимо отметить функционирование с его участием двух типов патосистемы: локально-раневой (в терминологии автора) и множественно-локальной (по Davis,1936).
Локально-раневая патосистема — следствие сопряженного развития видов, когда в результате питания фитофагов (онтогенетически приуроченного к питанию апикальной, интеркалярной и эмбриональной меристемами) облегчается проникновение, реализация агрессивности и завершение цикла развития облигатного паразита U. maydis. Она функционирует одновременно с патосистемой растение-хозяин — патоген, но важной характеристикой последней является множественность очагов распространения мицелия по направлению к меристематическим тканям, но на незначительное расстояние, частой локализацией грибницы в пазушных почках без видимых проявлений болезни. Такой характер распределения мицелия U. maydis по растению известен как множественно-локальный; локально-раневая патосистема, характеризующаяся проявлением галлов, учитывается при мониторинге распространения головни и олицетворяет объем успешных заражений растений определенного генотипа в природе, который намного меньше числа очагов множественно-локальной (скрытой) инфекции.
Описанные выше особенности отношений в системе растение-хозяин — фитофаги — патоген характеризуют моноциклический тип болезни, эпифитотийных проявлений которой не отмечено.
Семенами возбудитель не передается, их заспорение телиоспорами не приводит к возникновению болезни (в противоположность пыльной головне). Протравливание семян защищает проростки от широкого круга патогенов, в том числе от переноса телиоспор U. maydis в новые регионы. Включение в последние годы в системы защиты посевов зерновой кукурузы фунгицидов с целью снижения поражённости пузырчатой головней початков носит скорее превентивный характер, не связанный с прогнозируемым и экономически просчитанным недобором урожая. Такие обработки вероятно целесообразны в перечне приемов защиты сахарной кукурузы при прогнозируемо высокой численности вредителей початков, где недобор урожая от вредителей и головни выше, чем на обычной кукурузе.
Приводимые в научной, учебной научно-популярной и специальной литературе последнего десятилетия рекомендации (подобные тем, что процитированы в таблице 1) сформулированы на основе недостаточно четких представлений об этиологии болезни и её патогенезе.
Сопряженная патосистема (Zea mays — вредители (шведская муха, кукурузный мотылек, хлопковая совка) — U. maydis) является коэволюционно сложившейся, и в таком её значении дополняет базовую парадигму применительно к практической селекции на групповую и комплексную устойчивость к вредным организмам, её анализ позволяет оптимизировать разработку экологизированной системы защиты растений от фитофагов и патогенов. Она характеризуется проявлением у кукурузы эволюционно стабильной нерасоспецифической устойчивости, сохранение которой обеспечено правильно выбранной в начале ХХ века стратегией отбора. Сохранение этой устойчивости предполагает более широкий мониторинг раневых повреждений как показателя первичного выбора определенного генотипа растений (осуществляемого фитофагами), как природного фона, на котором облегчается выявление наименее повреждаемых и поражаемых селекционных образцов, как прогнозируемого и в значительной мере управляемого фактора в биологизации системы защиты кукурузы.
При составлении материала, мы использовали следующие статьи:




